생물체의 복잡한 세포 및 생리 학적 과정은 예를 들어 서식지에 대한 동물 또는 식물의 적응성을 보장하기 위해 분자 수준에서 미세 조정 된 조절이 필요합니다. 이를 위해 세포 전달, 신진 대사 또는 세포 분열과 같은 과정에 개입하는 수많은 분자가 있습니다. 이 분자 중 하나는 단백질입니다 칼 모듈 린칼슘의 도움으로 다른 많은 생물학적 활성 단백질의 기능에 영향을 미칩니다.
칼 모둘 린이 란?
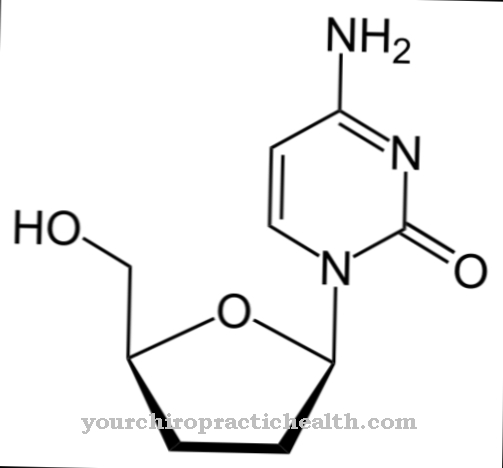
칼 모듈 린은 칼슘 이온을 결합하는 세포 내 조절 단백질입니다. 그 구조로 인해 EF-hand 단백질 그룹에 속합니다. 148 개의 아미노산으로 구성되어 있고 길이가 6.5nm 인 칼 모둘 린의 모양은 아령과 비슷합니다. 이 단백질 분자의 분자량은 약 17kDa입니다.
세포 내 신호 전달의 생물학적 기능으로 인해 칼 모둘 린은 두 번째 메신저, 즉 효소 적으로 활성이 아닌 이차 메신저 물질로 분류 될 수도 있습니다. 단백질의 2 개의 구형 도메인에는 1.1nm 거리에 2 개의 나선형 루프 나선형 모티프가 있으며 총 4 개의 칼슘 이온이 결합 될 수 있습니다. 이 구조를 EF 손이라고합니다. EF- 손 구조는 칼 모둘 린의 반 평행 베타 시트 사이의 수소 결합으로 연결됩니다.
기능, 효과 및 작업
칼 모듈 린이 활성화 되려면 분자 당 3-4 개의 결합 된 칼슘 이온이 필요합니다. 활성화 된 상태에서 형성된 칼슘-칼 모듈 린 복합체는 다양한 기능을 가진 수많은 수용체, 효소 및 이온 채널의 조절에 관여합니다. 조절 된 효소로는 면역 반응을 조절하는 데 중요한 역할을하는 포스파타제 칼시 뉴린과 NO를 생성하는 내피 산화 질소 신타 제 (eNOS)가 있습니다. 혈관.
낮은 칼슘 농도에서 adenylate cyclase (AC)는 높은 칼슘 농도에서 활성화되고 반면에 효소 대응 물인 PDE (phosphodiesterase)는 활성화됩니다. 이러한 방식으로 조절 메커니즘의 연대순 순서가 달성됩니다. 처음에는 AC가 순환 AMP (cAMP) 생산을 통해 이동중인 신호 경로를 설정하고 나중에는 cAMP 저하를 통해 상대 PDE에 의해 다시 꺼집니다. 그러나, 아래에서보다 상세히 설명 할 CaM 키나제 II 또는 미오신 경쇄 키나제 (MLCK)와 같은 단백질 키나제에 대한 칼 모둘 린의 조절 효과는 특히 알려져있다.
CAMKII는 인산 잔기를 다양한 단백질에 결합하여 에너지 대사, 이온 투과성 및 세포에서 신경 전달 물질의 방출에 영향을 미칠 수 있습니다. CAMKII는 특히 뇌의 고농도에서 발견되며, 여기서 신경 가소성, 즉 모든 학습 과정. 그러나 칼 모둘 린은 또한 운동 과정에 없어서는 안될 필수 성분입니다. 휴식시 근육 세포의 칼슘 이온 농도는 매우 낮으므로 칼 모둘 린은 비활성 상태입니다. 그러나 근육 세포가 흥분되면 칼슘이 세포 혈장으로 흘러 들어가고 보조 인자로서 칼 모둘 린의 4 개의 결합 부위를 차지합니다.
이것은 이제 미오신 경쇄 키나아제를 활성화시켜 세포의 수축 섬유를 이동시켜 근육 수축을 가능하게합니다. 칼 모둘 린의 영향을받는 다른 잘 알려지지 않은 효소는 구 아닐 레이트 사이 클라 제, Ca-Mg-ATPase 및 포스 포 리파제 A2입니다.
교육, 발생, 속성 및 최적의 가치
칼 모듈 린은 모든 식물, 동물, 균류 및 아메바 이드 생물 군을 포함하는 모든 진핵 생물에서 발생합니다. 이 유기체의 칼 모둘 린 분자는 일반적으로 비교적 유사한 방식으로 구조화되어 있기 때문에 초기 단계에서 출현 한 진화 적으로 매우 오래된 단백질이라고 가정 할 수 있습니다.
일반적으로 칼 모둘 린은 세포의 혈장에 비교적 많은 양으로 존재합니다. 예를 들어 신경 세포의 세포질에서 일반적인 농도는 약 30-50 µM입니다. 0.03-0.05 mol / L. 단백질은 CALM 유전자를 사용하여 전사 및 번역 중에 형성되며, 그 중 CALM-1, CALM-2 및 CALM-3이라고하는 현재까지 알려진 세 개의 대립 유전자가 있습니다.
질병 및 장애
칼 모둘 린에 대한 억제 효과를 가질 수있는 일부 화학 물질이 있으므로 칼 모둘 린 억제제로 알려져 있습니다. 대부분의 경우 억제 효과는 칼슘을 세포 밖으로 운반하여 칼 모둘 린에서 제거하여 비활성 상태로만 존재한다는 사실에 기반합니다.
이러한 억제 물질에는 예를 들어 W-7이 포함됩니다. 또한 일부 향정신성 페 노티 아진 약물은 칼 모둘 린을 억제합니다. 칼 모둘 린의 조절 기능만큼 광범위하고 단백질이 더 이상 보조 인자 칼슘에 의해 활성화 될 수없고 조절 된 표적 효소 자체가 덜 활성화 될 때 상상할 수있는 결함과 장애가 매우 다양합니다. 예를 들어, CAMKII의 부적절한 활성화는 학습 과정의 기초를 형성하는 신경 가소성의 제한으로 이어질 수 있습니다.
MLCK의 활성화가 감소하면 근육 수축이 손상되어 운동 장애가 발생할 수 있습니다. 칼 모둘 린 결핍으로 인한 칼시 뉴린 효소의 활성화가 적 으면 신체의 면역 반응에 영향을 미치고 eNO의 활성화가 적 으면 NO 농도가 낮아집니다. 후자는 주로 산화 질소가 원하지 않는 혈액 응고를 방지하고 더 나은 혈액 순환을 위해 혈관을 확장시키는 문제를 일으 킵니다. 그러나이 시점에서 칼슘 센서 Frequenin이 특정 조건에서 칼 모둘 린의 생물학적 기능을 대신하여 분자를 대체 할 수 있다는 점도 언급해야합니다.





.jpg)